Водоподъемные установки | Mensh.ru
Опубликовано mensh — ср, 09/14/2011 — 09:34
Инженерные системы
Для индивидуальных систем водоснабжения следует применять автоматизированные водоподъёмные установки, включающие в себя насос, регулирующую ёмкость и приборы автоматического регулирования, при которых насос периодически подаёт или прекращает подачу воды в регулирующую ёмкость в зависимости от уровня воды в безнапорном (открытом) баке или давления в напорном гидропневматическом баке.
Работа водоподъёмной установки характеризуется частотой включения насоса в единицу времени, зависящей от регулирующего объёма бака.
Безнапорный (открытый) бак размещается в высшей точке системы на отметке, обеспечивающей необходимый напор в системе.
В напорном гидропневмобаке необходимый напор создаётся давлением сжатого воздуха, передающемся на воду через эластичную мембрану.

 Регулирующий объём ёмкости W, м3, надлежит определять по формуле
W = qhrsp/4n, (2)
где qhrsp —производительность насоса, м3/ч;
n —допустимое число включений насоса в час, принимаемое для установок:
Регулирующий объём ёмкости W, м3, надлежит определять по формуле
W = qhrsp/4n, (2)
где qhrsp —производительность насоса, м3/ч;
n —допустимое число включений насоса в час, принимаемое для установок:- с безнапорными баками —до 4;
- с гидропневмобаками —до 10.
- 0,8 —для установок, работающих с подпором;
- 0,75 —для установок с напором до 50 м;
- B —коэффициент запаса вместимости бака, принимаемый 1,2–1,3.
Высота расположения безнапорного бака и минимальное давление в гидропневматическом баке должны обеспечивать необходимый напор воды перед водоразборной арматурой.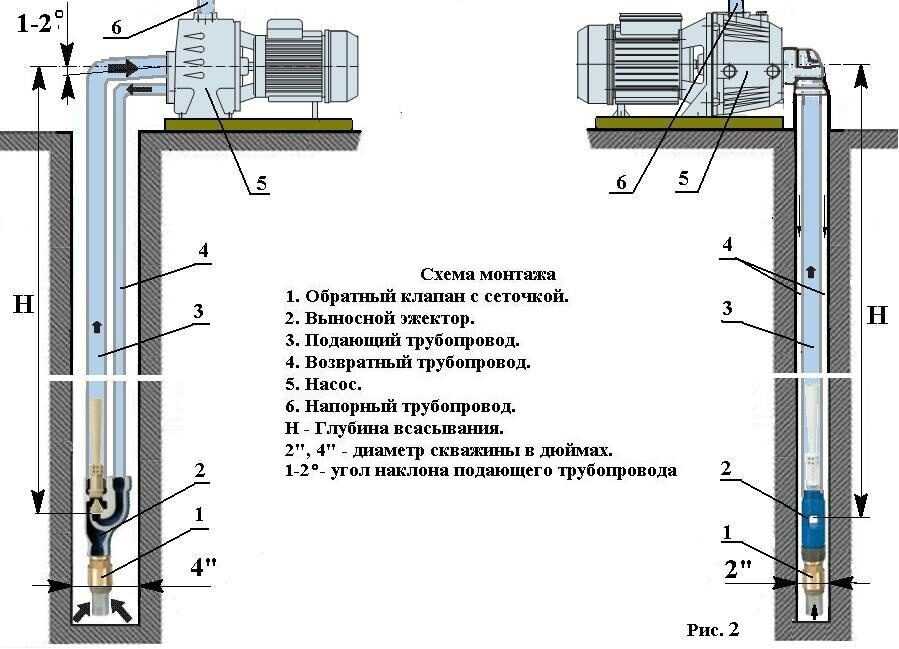

Устройство и работа
Установка с гидроаккумулятором ВУ-16-28 (рис. 4.2) состоит из: гидроаккумулятора — 4; насосного агрегата — 5; станции управления — 20; системы автоматики – 13,14,15,16,17,20; водопроводной арматуры – 6,7,9,12.
Рис. 4.2. Общий вид водоподъёмной установки ВУ-16-28А.
1
– помещение подземное: 2 – люк; 3 – труба
вентиляционная; 4 – гидроаккумулятор;
5
– насос с электродвигателем; 6 –
трубопровод всасывающий; 7 – клапан
приемный; 8 – скважина; 9 – магистраль
водоразборная; 10 – кран трехходовой;
11 – кабель электрический; 12 – трубопровод
напорный; 13 – трубка реле давления; 14 –
реле давления; 15 –диафрагма; 16 – концевой
выключатель; 17 – ограничитель хода
диафрагмы; 18 – штуцер для закачки
воздуха; 19 – манометр; 20 – станция
управления.
Установка работает в автоматическом режиме. Управление режимом работы осуществляется станцией, управления — 20 и датчиком — реле давления — 14. Вода из водоисточника – 8, насосом – 5, через трехходовой кран — 10 подаётся по трубопроводу — 12 к гидроаккумулятору — 4 и в водоразборную магистраль — 9. Давление в трубопроводе — 12 и водоразборной магистрали — 9 изменяется в зависимости от расхода воды потребителем и достигает максимума
 По
мере потребления воды потребителем
давление в трубопроводе снижается до Р1,
реле давления подаёт сигнал станции
управления на включение электродвигателя
насоса. Цикл повторяется.
По
мере потребления воды потребителем
давление в трубопроводе снижается до Р1,
реле давления подаёт сигнал станции
управления на включение электродвигателя
насоса. Цикл повторяется.Давление включения и выключения устанавливается по расчетным данным потребителем на датчик-реле давления — 14. Давление воздуха и воды контролируется манометром — 19.
В
случае утечки воздуха из гидроаккумулятора
во время работы водокачки воздушная
подушка в верхней камере уменьшается,
в результате этого диафрагма – 18, под
давлением воды, создаваемым насосом,
будет подниматься выше нормального
рабочего положения, что может привести
к порыву последней. Для того, чтобы
исключить создавшуюся аварийную
ситуацию, гидроаккумулятор оснащен
ограничителем хода диафрагмы — 17.
Диафрагма, поднимаясь выше нормального
рабочего положения, перемещает
ограничитель хода диафрагмы, ось качания
которого находится на уровне стенки
гидроаккумулятора.
Сколько энергии тратят растения на подъем воды из почвы? • Earth.com
09-08-2022
АвторAndrei Ionescu
Штатный сотрудник Earth.com
Каждый день примерно один квадриллион галлонов воды бесшумно перекачивается деревьями от их корней к верхушкам деревьев – поразительный подвиг, совершенный используя только солнечный свет. Хотя ученым давно известно, что для подъема всей этой жидкости требуется огромное количество энергии, но каково именно это количество, оставалось загадкой.
Теперь группа исследователей из Калифорнийского университета в Санта-Барбаре впервые подсчитала огромное количество энергии, используемой растениями для перемещения воды через их ксилему — тип транспортной ткани сосудистых растений — к их листьям. Их анализ показал, что в среднем он составляет до 14,2% энергии, которую растения собирают в процессе фотосинтеза. Это количество сравнимо с производством гидроэнергии всего человечества.
«Требуется сила, чтобы поднять воду по ксилеме дерева. Это требует энергии. Мы подсчитываем, сколько это энергии», — сказал ведущий автор исследования Грегори Кетин, научный сотрудник в области экологии из Калифорнийского университета в Санта-Барбаре. Эта энергия является излишком того, что растения производят посредством фотосинтеза. «Энергия пассивно извлекается из окружающей среды, просто через структуру дерева».
Сложные растения совершают этот удивительный подвиг с помощью сосудистой системы, в которой трубки, называемые ксилемой, доставляют воду от корней к листьям, а другой тип трубок, известный как флоэма, транспортирует сахар, вырабатываемый в листьях, к остальным частям растения. «Сосудистые растения, развивающие ксилему, — это огромное дело, которое позволило деревьям существовать», — объяснил доктор Кетин.
В отличие от систем кровообращения животных, системы кровообращения растений открыты и питаются пассивно, при солнечном свете вода испаряется, которая затем выходит из пор в листья и создает отрицательное давление, которое поднимает воду под ним посредством процесса, называемого «транспирация». ». В отличие от фотосинтеза, энергию транспирации не нужно обрабатывать перед использованием.
». В отличие от фотосинтеза, энергию транспирации не нужно обрабатывать перед использованием.
Ученые объединили глобальную базу данных о проводимости растений с математическими моделями подъема сока, чтобы рассчитать, сколько энергии заводы тратят на перекачку воды. Они обнаружили, что леса Земли потребляют около 9,4 петаватт-часов в год — количество, аналогичное мировому производству гидроэлектроэнергии. Удивительно, но небольшая часть этой энергии расходуется на борьбу с гравитацией, большая ее часть идет на преодоление сопротивления стеблей растений.
Поскольку растения сильно различаются по степени приспособленности их стеблей к потоку воды, все еще существует некоторая неопределенность в отношении того, сколько энергии производят те или иные растения. «Я думаю, что эта неопределенность подчеркивает, что мы еще многого не знаем о биогеографии устойчивости растений (и, в меньшей степени, транспирации). Это хорошая мотивация для продолжения исследований в этих областях», — заключил доктор Кетин.
Исследование опубликовано в Журнале геофизических исследований: Биогеонауки .
—
по Andrei Ionescu , Earth.com Управляющий писатель
Связанные новости
Новости. США
Политика конфиденциальности
Условия использования
Карта сайта
Персонал
Earth.com
News
Видео
Изображения
Earthedia
Действие
Магазин
Earthedia
Encyclopedia
.
Последние новости
Окружающая среда
Образ жизни
Голоса
Животные
Растения
СВЯЖИТЕСЬ
Свяжитесь с нами
© 2023 Earth.com
Все права защищены
Гидравлический подъем и использование воды растениями: влияние на водный баланс, производительность и взаимодействие между растениями
Allen EB, Allen MF (1986) Водные отношения ксеровых злаков в поле: взаимодействие микориз и конкуренция. Новый фитол 104:559–571
Новый фитол 104:559–571
Google Scholar
Baker JH, Bavel CHM van (1986) Устойчивость корней растений к потере воды. Агрон Дж. 78: 641–644
Google Scholar
Бельский А.Дж., Амундсон Р.Г., Даксбери Дж.М., Риха С.Дж., Али А.Р., Мвонга С.М. (1989) Влияние деревьев на их физическую, химическую и биологическую среду в полузасушливой саванне в Кении. J Appl Ecol 26:1005–1024
Google Scholar
Biswell HH (1935) Влияние окружающей среды на корневую систему некоторых лиственных лесных деревьев. Бот Газ 96: 676–708
Google Scholar
Bormann FH (1957) Перенос влаги между растениями через переплетение корневых систем. Завод Физиол 32:48–55
Google Scholar
Коробка ЭО (1981) Макроклимат и растительная форма. Dr W Junk Publ Гаага
Dr W Junk Publ Гаага
Google Scholar
Boyer JS (1985) Водный транспорт. Энн Рев Плант Физиол 36:473–516
Google Scholar
Boyer JS (1982) Продуктивность растений и окружающая среда. Наука 218:443–448
Google Scholar
Браун Р.В., Бартос Д.Л. (1982) Калибровочная модель для психрометров с термопарой Пельтье с экраном. Исследовательский документ Лесной службы Министерства сельского хозяйства США INT-293, Огден, Юта, стр. 155
Колдуэлл М.М. (1985) Холодные пустыни. В: Шабо Б.Ф., Муни Х.А. (ред.) Физиологическая экология растительных сообществ Северной Америки. Чепмен и Холл, Нью-Йорк. стр. 198–212
Google Scholar
Caldwell MM, Richards JH (1989) Гидравлический подъемник: отток воды от верхних корней повышает эффективность поглощения воды глубокими корнями. Экология 79:1–5
Экология 79:1–5
Google Scholar
Coleman ML, Shepard TJ, Durham JJ, Rouse JE, Moore GR (1982) Восстановление воды цинком для анализа изотопов водорода. Анальная химия 54:993–995
Google Scholar
Корак С.Дж., Блевинс Д.Г., Палларди С.Г. (1987) Перенос воды в ассоциации люцерны/кукурузы. Завод Физиол 84:582–586
Google Scholar
Доусон Т.Е. (1993a) Источники воды растений, определенные по изотопному составу ксилемы и воды: перспективы конкуренции растений, распределения и водных отношений. В: Элерингер Дж. Р., Холл А. Е., Фаркуар Г. Д. (ред.) Стабильные изотопы и отношения углерода и воды в растениях. Академик Пресс, Сан-Диего. стр. 465–496
Google Scholar
Доусон Т.Е. (1993b) Водный баланс лесов. Тренды Ecol Evol 8:120–121
Google Scholar
Доусон Т.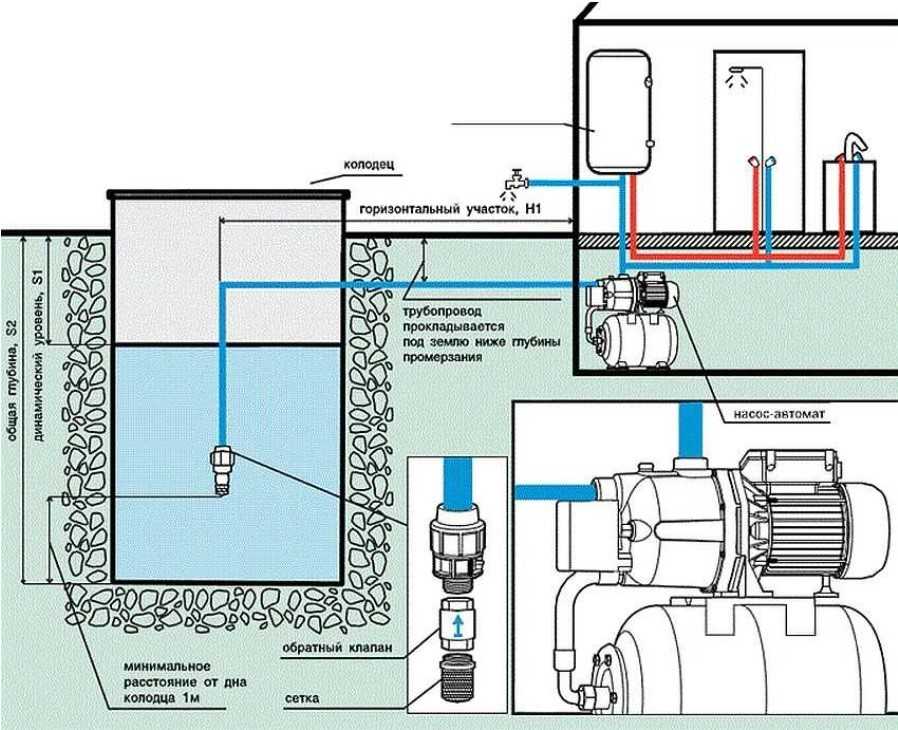 Е., Элерингер Дж.Р. (1991) Прибрежные деревья, не использующие речную воду. Природа 350:335–337
Е., Элерингер Дж.Р. (1991) Прибрежные деревья, не использующие речную воду. Природа 350:335–337
Google Scholar
Доусон Т.Е., Элерингер Дж.Р. (1993a) Гендерная физиология, дискриминация изотопов углерода и распределение среды обитания у боксэлдера, 9 лет.0202 Клен черный . Экология 74:798–815
Google Scholar
Доусон Т.Э., Элерингер Дж.Р. (1993b) Обогащение изотопами воды в «древесных» тканях растений: значение для источника воды растениями, водопоглощение и другие исследования, в которых используется стабильный изотопный состав целлюлозы. Геохим Космохим Акта 57:3487–3492
Google Scholar
Дирксен К., Раатс ПАК (1985) Поглощение и выделение воды корнями люцерны. Агрон Дж. 77: 621–626
Google Scholar
Ehleringer JR, Dawson TE (1992) Поглощение воды растениями: перспективы стабильных изотопов. Plant Cell Envir 15:1073–1082
Plant Cell Envir 15:1073–1082
Google Scholar
Ehleringer JR, Osmond CB (1989) Стабильные изотопы. В: Pearcy RW, Ehleringer JR, Mooney HA, Rundel PW (eds) Физиологическая экология растений. Полевые методы и аппаратура. стр. 281–300 Chapman & Hall, Publishers, Лондон, Англия 9.0003
Google Scholar
Eissenstat DM, Caldwell MM (1988) Конкурентоспособность связана со скоростью извлечения воды. Полевое исследование двух дерновинных трав в засушливых районах. Экология 75:1–7
Google Scholar
Fayle DCF (1965) Привычка укоренения сахарного клена и желтой березы. Dept For Canada Publ 1120, Ottawa
Griffiths B, Robinson D (1992) Минерализация азота, вызванная корнями: модель баланса азота. Растительная почва 139: 253–263
Google Scholar
Hansen SE, Dickson RE (1979) Перенос воды и минеральных питательных веществ между корневыми системами молоди Populus . Лесные науки 24: 247–252
Лесные науки 24: 247–252
Google Scholar
Дженсен Р.Е., Тейлор С.А., Вибе Х.Х. (1961) Отрицательный транспорт и сопротивление потоку воды через растения. Завод Физиол 36:633–638
Google Scholar
Жоффр Р., Рамбал С. (1993) Как лесной покров влияет на водный баланс средиземноморских пастбищных угодий. Экология 74:570–582
Google Scholar
Жоффр Р., Рамбал С. (1988) Улучшение почвенной влаги деревьями на пастбищах южной Испании. Acta Oecologia 9:405–422
Google Scholar
Kramer PJ (1983) Водные отношения растений. Академик Пресс, Нью-Йорк
Google Scholar
Купперс М., Нилс Т.Ф., Купперс Б.Л., Суон А.Г., Майерс Б.А. (1987) Характеристики гидравлического потока в лигнобугорчатом молотке Eucalyptus behriana F. Muell. в поле. Plant Cell Envir 10: 27–37
Muell. в поле. Plant Cell Envir 10: 27–37
Google Scholar
Мазор Е. (1991) Прикладная химическая и изотопная гидрология подземных вод. Издательство Открытого университета. Великобритания
Google Scholar
McCully ME, Canny MJ (1988) Пути и процессы движения воды и питательных веществ в корнях. Растительная почва 111:159–170
Google Scholar
Molz FJ, Peterson CM (1976) Перенос воды от корней к почве. Агрон Дж. 68:901–904
Google Scholar
Муни Х.А., Гулмон С.Л., Рундел П.В., Элерингер Дж.Р. (1980) Дальнейшие наблюдения о водных отношениях Prosopis tamarungo северной пустыни Атакама. Экология 44:177–180
Google Scholar
Нили Дж. А. (1965) Обследование почвы в округе Томпкинс, штат Нью-Йорк. Отчет службы охраны почв Министерства сельского хозяйства США. Типография правительства США, Вашингтон, округ Колумбия
Отчет службы охраны почв Министерства сельского хозяйства США. Типография правительства США, Вашингтон, округ Колумбия
Google Scholar
Nobel PS, Cui M (1992) Прогнозирование и измерение проводимости водяного пара в зазорах для корней, расположенных концентрически и эксцентрично в воздушных зазорах. Растения и почва 145:157–166
Google Scholar
Nobel PS, Sanderson J (1984) Rectifer-подобная деятельность корней двух пустынных суккулентов. J Exp Bot 35: 727–737
Google Scholar
Oertli JJ (1991) Транспорт воды в ризосфере и корнях. В: Вайсель Ю., Эшель А. (ред.) Корни растений: скрытая половина. Марсель Деккер, Нью-Йорк. стр. 559–587
Google Scholar
Passioura JB (1988) Транспорт воды в корни и к корням. Ann Rev Plant Physiol Plant Molec Biol 39:245–265
Google Scholar
Ричардс Дж. Х., Колдуэлл М. М. (1987) Гидравлический подъем: постоянный ночной перенос воды между слоями почвы с помощью корней Artemisia tridentata . Экология 73:486–489
Х., Колдуэлл М. М. (1987) Гидравлический подъем: постоянный ночной перенос воды между слоями почвы с помощью корней Artemisia tridentata . Экология 73:486–489
Google Scholar
Шипперс Б., Шрот М.Н., Хильдебранд Д.К. (1967) Эманация воды из подземных частей растений. Пант почва 27:81–91
Google Scholar
Schulze E-D (1986) Обмен углекислым газом и водяным паром в ответ на засуху в атмосфере и почве. Ann Rev Plant Physiol 37:247–274
Google Scholar
Schulze E-D, Robichaux RH, Grace J, Rundel PW, Ehlerinnger JR (1987) Водный баланс растений. Бионаука 37: 30–37
Google Scholar
Schwenke H, Wagner E (1992) Новая концепция корневой экссудации. Envir Plant Cell 15:289–299
Google Scholar
Шон М. Г.Т., Флад А.В. (1980) Исследования поглощения и потери воды корнями ячменя в связи с изменениями сопротивления корней. J Exp Бот 31:1147–1159
Г.Т., Флад А.В. (1980) Исследования поглощения и потери воды корнями ячменя в связи с изменениями сопротивления корней. J Exp Бот 31:1147–1159
Google Scholar
Стивенсон, Н.Л. (1990) Климатический контроль за распространением растительности: роль водного баланса. Ам Нат 135:649–670
Google Scholar
Thorburn PJ, Mensforth LJ (1993) Извлечение воды из медикаго сатива для анализа стабильных изотопов воды. Comm Soil Sci Plant Anal 24: 549–557
Google Scholar
Торберн П.Дж., Уокер Г.Р. (1993) Источник воды, испаренной Eucalyptus camaldulensis: почва, грунтовые воды или ручьи? В: Элерингер Дж. Р., Холл А. Е., Фаркуар Г. Д. (ред.) Стабильные изотопы и отношения углерода и воды в растениях. Академик Пресс, Сан-Диего. стр. 511–527
Google Scholar
Tyree MT, Ewers FW (1991) Гидравлическая архитектура деревьев и других древесных растений.